
© Dario J Lagana - Fondazione MAXXI Rome
Spiders are not insects in the taxonomic sense, but belong to a broader group of invertebrates; insects, in fact, make up the main diet of spiders. However, prey and predator population dynamics can be paradoxical, existing in synergistic relationship with each other. Spiders and insects need one another; or, to look at it differently, both spiders and insects play a vital role in the biological systems that support life on Earth.
The ‘Animal Internet’ describes a proliferation of collaboratively generated GPS and other geotagged online data that assists scientists in mapping the distribution, movements and interactions of various animal populations. For example, the website and app inaturalist allows users to upload species data and locations, and/or identify unknown plant and animal species. In turn, data from the Animal Internet can be used to better understand patterns of extinction.
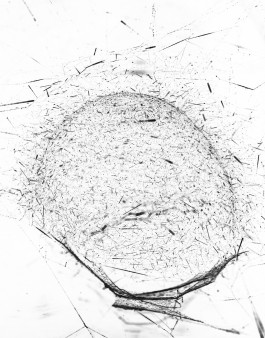
In 2019, Studio Tomás Saraceno is mapping the synanthropic spider/web ecologies of Venice, both inside its anthropogenic architectures, and in the areas less touched by human hands.
On a mapping exercise undertaken by Tomás Saraceno and members of the Spider/Web Research Group in early 2019, we encountered a number of spider/web typologies in the grounds of the Venice Biennale, as marked in the map below. We invite you - through processes of becoming sensitive and attuned to the multispecies ecologies in which we are embedded - to add to this map - by uploading photos of the spider/webs that you encounter.

Collage with Matthäus Merian, 1650 and Jacopo de’ Barbari Map of Venice, 1500.

Sheet webs
Bathyphantes gracilis
Centromerus sylvaticus
Diplocephalus connectens
Erigone dentipalpis
Lepthyphantes istrianus
Lepthyphantes tenuis
Mecopisthes nicaeensis
Meioneta mollis
Meioneta rurestris
Space webs
Amaurobius erberi
Haplodrassus dalmatensis
Microctenonyx subitaneus
Pachygnatha degeeri
Pelecopsis krausi
Prinerigone vagans
Tiso vagans
Trichopterna cito
Steatoda phalerata

Orb webs
Mangora acalypha
Zilla diodia

In between the elm branches and in the hands of Aracne, worlds are folded in the Dialettica. They are told to announce clear skies, tie up a lion if they unite or that, as words, they could be both shelters and traps. Nothing makes itself alone: ask the oracles how many multitudes might contain the cosmic web.
In May 2018, Studio Tomás Saraceno made a survey of the spider/webs of Palais de Tokyo in collaboration with Christine Rollard from Muséum national d'Histoire naturelle (Paris). More than 500 spider/webs were found scattered throughout the gallery, in spaces least frequented or noticed: air conditioning vents, loading bays, and ceiling rafters. The most abundant spider/webs in the Palais de Tokyo are the dome-like webs built by Holocnemus pluchei. These spiders are synanthropes: species that thrive in human-modified environments, and whose ecologies are interdependently entangled with our own. There is an implied bond in sharing a house, a habitat, an ecosystem. Whether or not it is enacted in our social relations, our ecological entanglement demands that we find new sensitivities for collective living with the creatures and objects with whom we share this air.
Following the theories of the biologist Jakob von Uexküll, what we consider habitat is an act of perception: just as our milieu conditions or ‘captures’ us, it is simultaneously constructed by our perception of it. The spider/web’s perceptual faculties are different from our own; what it perceives as the shape of the world is estranged from us. Even if we share this room, it is markedly different for each of us. By paying at-tent(s)ion, we create worlds: bestowing meaning on our habitat, giving it the power to affect us. Perhaps we need to close our eyes, and stretch our arms in blindness, to begin to perceive the world as the spider/web does.
The habitats that envelop different species are not fixed. Paying at-tent(s)ion to the dynamic ways in which our own habitats take shape allows us to speculate on the kinds of practices for interspecies attunement (even multispecies jamming sessions) that might open up connections between worlds, transforming them and us in the process. If we imagine ourselves as being with worlds, rather than being in them—we might develop better practices for sharing them with other species.
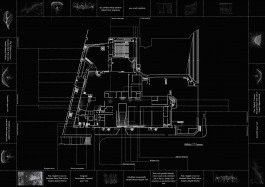
Tomás Saraceno, Spider/Web map of the Palais de Tokyo, Paris, 2018

© Andrea Rossetti, 2017

Since 2005, Tomás Saraceno has been articulating a shift in focus to tell stories from the perspective of the web—which is not separate to the spider, but a material extension of its senses and thoughts, an assemblage we think of as a spider/web. Museological collections tend to focus on the spider in isolation of its web, rather than tracing the material architectures that connect the spider/web to the world, and that tell different stories about evolution and adaptation. In response to this, and in collaboration with people and institutions across knowledge fields, Studio Tomás Saraceno is building the only existing archive of three-dimensional spider/webs—a shared resource for scientific and artistic inquiry, articulating a new classification that emerges from the web itself.
Spider/webs are dynamic architectures, not static objects—the spider continually adjusts, destroys and re-makes its web in response to shifting cues in its milieu. The webs documented months ago may not exist anymore in the places marked on this map. Like the spider/web, a map is not static or fixed, but open and emergent, and constitutes an invitation to explore. As you search, you will discover new webs, new habitats and perhaps even new species, as a living assemblage of spider/webs exists above our heads.
The irregular, three-dimensional space web built by Holocnemus pluchei often includes a dome-shaped structure: an element more likely to be present in the webs of gestating spiders. Most of the time, however, this long-legged, ‘kleptoparasitic’ spider can be found on the peripheries of the colonial webs of another Mediterranean spider, Cyrtophora citricola, whose web provides a ready source of food for H. pluchei. The C. citricola web shares some features with H. pluchei’s web: a horizontal orbweb with a very fine rectangular mesh, its hub suspended and elevated, giving it the shape of a tent, rather than the dome shape visible in H. pluchei webs. H. pluchei steals food, undetected, from the complex C. citricola web, while generally avoiding aggressive behavior. The kleptobiotic behaviour of spiders from the Pholcidae family is facilitated by their extremely long and delicate legs, allowing them slow and stealthy movements whose vibrations are nearly imperceptible to the host web spider. When the web is occupied and controlled by its original builder (C. citricola, or other species), H. pluchei stays in the periphery of the web and feeds on small arthropods, not building a web of its own. Occasionally, when there is a lack of prey in the web periphery, H. pluchei approaches the center of the web and steals prey that has been hunted, digested and guarded by C. citricola - sometimes even touching the host with its forelegs without provoking attack. When the original host dies or leaves the web, H. pluchei repurposes it for the construction of its own web.

Like most spiders, Eratigena atrica uses its web as a hypersensitive sensory system for sending and receiving vibrations, and through which it builds a dynamic image of its environment. Appended to this spider’s flat and dense sheet web is a funnel-shaped retreat (often located in the corner of the room in which the web is built), where the spider hides. From this silken retreat, the spider modulates the web in response to the vibrations that it senses, occasionally leaving the funnel to catch prey. The spider’s leg tip is very well-adapted to locomotion on its web: making Eratigena and Tegenaria species the fastest moving spiders in the world. Outside the web these spiders are less agile, and unable to ascend smooth surfaces - which is why we often find them trapped in bathtubs and sinks. Despite their eight eyes, most web-building spiders have very poor vision. The emerging research fields of biotremology (the study of substrate-borne signals in animals) and cognitive science have argued that the spider web can be thought of as an extension of the spider’s senses - and even, some have argued, of the spider’s cognitive system. The spider web is not only a medium to receive signals from trapped prey, but also a communication channel used for intraspecific signaling, mainly during courtship. In 1880, C.V. Boys published a paper in Nature describing the reactions of a web-building spider to the vibrations of a tuning fork ('The Influence of a Tuning-Fork on the Garden Spider'). Since then, this technique of using a vibrating source - including even sonic toothbrushes - to induce a response from spiders is common in arachnological research practice.
The flat brushed sheet web and its domestic house spider inhabitant, Tegenaria domestica, have a longstanding synanthropic relationship with humans. Over thousands of years, these spider/webs have been cohabiting, adapting and mutually evolving alongside humans and human habitats. Initially built in caves and cracks in rocks, the web has been adapted to structures produced by anthropocenic development: houses, buildings, even shopping malls, which have become the preferred habitat. Reportedly the most common house spider in England until the 19th century, T. domestica later relinquished this title to its larger, close relative, Eratigena atrica (the giant house spider). Tegenaria (and Eratigena) spiders build dense sheet webs, commonly found in the quiet corners of our homes. The sheet web differs from the orb web - whose geometry provides obvious spatial cues or ‘landmarks’ to the spider for orientation. In the sheet web, the spider may be more reliant on vibrational cues, as well as external cues such as light position, kinaesthetic or barostatic signals, to orient itself within the web. In contrast to most Central European spiders, these species can live for several years. T. domestica also holds the world record of the fastest moving spiders: the special claws on their legs allow them move very quickly and nimbly along the threads of their webs.
La Fausse veuve noire ou Steatode domestique
This ‘gumfoot’ or space web is composed of many threads expanding in different directions. It is built by the so-called 'false black widow', Steatoda grossa, who shares many similarities with Latrodectus mactans, the true black widow spider, and its web. The lower threads of the web that are attached to the soil are under tension. When a small creature encounters and becomes entangled within these tension threads, the threads detach from the soil and ‘flip’ the prey into the web. This kind of web is very effective, allowing the spider to catch scorpions and other large prey. The dynamics of this snare also make it difficult for potential mates to approach one another. Male spiders therefore use specific vibrational signals to communicate with the female. If the female spider is not receptive, she will simply reject the male spider from the web. S. grossa belongs to the same Theridiidae family of cobweb-building spiders to which the true black widow belongs: a diverse and cosmopolitan group of arachnids. However, a bite from the false black widow - while similar to that of the true black widow - is less severe. Another key difference between the true and false black widows is in their mode of vibrational communication. The stridulating signals produced by S. grossa males to communicate with their mates are airborne sounds audible to humans. This sound is produced via friction that the spider generates between parts of its body, and elicits female receptivity during courtship. This form of stridulating communication is essential for intra-species recognition, and for the species’ reproductive process.

The small, mesh-like web built by Brigittea civica includes cribellate silk. Web-building spiders are loosely divided into two main groups, according to the kind of silk that they produce: cribellate and ecribellate. Cribellate spiders have a special organ (cribellum) which allows them to produce 'woolly silk' which has very fine fibers, a large surface structure, and is strongly adhesive. As a result, cribellate spiders do not need to ‘dot’ their silk with drops of adhesive glue - an action performed by ecribellate spiders to make their webs more sticky, and thus better able to catch prey. An advantage of cribellate silk is that it remains adhesive for longer periods, where the ‘glue’ used by ecribellate spiders can dry out over time. Originally a rare species from Southern Europe, B. civica is now frequently found in colonies on the facades of buildings, also in northern parts of Europe. Individual B. civica spiders (2 to 4 mm) spin small but dense and rather resistant or hardy webs, known to accumulate dust and dirt over time as a testament to their longevity. These webs are often perceived as an ‘aesthetic’ inconvenience: pest-control companies offer services to mechanically remove such webs, and in some cases also provide special pesticidal treatments to remove B. civica and its webs from the faces of the buildings it has colonised. Ironically, these spider webs are also responsible for catching and thus eradicating other ‘undesirable’ insects - such as mosquitoes - from these same buildings.
Studio Tomás Saraceno is located in the Rummelsburg district of Berlin, near the banks of the Rummelsburger See, fed by the River Spree. During much of the 19th and 20th centuries this area was devoted to industrial activity, also housing what was, for a time, Europe’s largest coal-fired power plant. Since the mid 1990s, the area has been undergoing a renaturation process, including the development of a green belt along the lake and river banks. Spider/webs have always been part of the multispecies Rummelsburg ecology; coexisting synanthropically with their human kin through the area’s industrial and postindustrial phases. A mapping exercise undertaken by Studio Tomás Saraceno in early 2019 uncovered ten taxonomic species distributed among over 350 spider/webs that had taken up residence in the corners, crevices and rafters of the main,three-storey building of Studio Tomás Saraceno. The spider/webs we encountered differered according to six major web typographies, outlined below. The most prevalent spider/web species in this habitat was Pholcus phalangioides and its irregular space web.


Pholcus phalangioides

Eratigena atrica
Amaurobius

Irregular, horizontally-oriented space web -- Pholcus phalangioides
Pholcus phalangioides - the Pholcid or cellar spider- builds an irregular ‘space web’ that is almost ubiquitous in different human-made environments. Built on a horizontal plane, these webs are often not clearly separated from neighbouring spider/webs, such that the inhabitants of proximal webs exist in a ‘friendly’ kind of coexistence.

Tangle web -- Parasteatoda species
The prefix ‘para’ is derived from Ancient Greek, meaning beside or near, and Parasteatoda spider/webs are found in constant proximity to humans. As such, these spiders are not usually aggressive, and may tolerate humans approaching their web. Their tangled webs are often found in secluded areas such as attics, in the spaces between loose walls and behind open doors.
Missing sector orb web -- Zygiella x-notata
Zygiella x-notata builds an orb web with a conspicuous ‘hole’, a missing sector in the upper corner of the web, in which the spider places its signal thread. This thread is connected to the hub or centre of the orb where the spider rests, delivering critical vibrational information.
Flat, slightly concave brushed sheet web with a funnel-shaped retreat -- Eratigena atrica/ Tegenaria domestica
Appended to this spider’s flat and dense sheet web is a funnel-shaped retreat (often located in the corner of the room in which the web is built), where the spider hides. From this silken retreat, the spider modulates the web in response to the vibrations that it senses, occasionally leaving the funnel to catch prey. The spider’s leg tip is very well-adapted to locomotion on its web: making Eratigena and Tegenaria species the fastest moving spiders in the world.

Orb web -- Araneus diadematus
The garden spider is a widespread and well-known orb-weaver, which builds its spiral, two-dimensional web during the early morning hours. These webs are usually large in relation to the spider, and more dense in the hub or centre of the orb. The spider will eat and digest its web before building a new one, making it a perfect recycler of material. By eating its web, the spider also gains additional protein nutrition via the pollen grains that were caught in the threads of the web.


Lace web made with cribellate silk -- Amaurobius species
These cribellate lace spider/ webs are often found in and around buildings, and are particularly prevalent near windows. The cribellate silk gives the spider/web a woolly appearance, sometimes with a shiny blue colour due to the friction of the light. The older lace spider/webs can appear dense, felted and gray.


Modified, horizontally oriented dome-shaped space web -- Holocnemus pluchei
The irregular, three-dimensional space web built by Holocnemus pluchei often includes a dome-shaped structure: an element more likely to be present in the webs of gestating spiders. Most of the time, however, this long-legged, ‘kleptoparasitic’ spider can be found on the peripheries of the colonial webs of another Mediterranean spider, Cyrtophora citricola, whose web provides a ready source of food for H. pluchei. The C. citricola web shares some features with H. pluchei’s web: a horizontal orbweb with a very fine rectangular mesh, its hub suspended and elevated, giving it the shape of a tent, rather than the dome shape visible in H. pluchei webs.

The diving bell spider (Argyroneta aquatica) lives almost completely underwater, in submerged air bubbles that she captures from the water’s surface, and then drags below, securing with a few threads of silk to form her diving bell. An air-breathing animal, the spider is not biologically equipped for underwater living: she uses oxygen trapped in the hairs that cover its body - giving the spider its mercurial appearance -, as well as the air contained within its underwater bell, similar to the diving suit we humans would use to breathe underwater, carrying our oxygen supplies. Over time, the oxygen content of the diving bell becomes depleted, at which point the spider returns to the surface to collect more air to replenish its underwater home.
A literal “amphibian”, Argyroneta aquatica has at least two different kinds of life, being able to live both on land or in water - she inspires us to become what Eben Kirksey would call “ontological amphibians” (Kirksey, 2016) being able to have two or more modes of existence, travelling across Umwelten or specific worlds.
From being on air, atmosphere-bound and terrestrial beings, how could we dive in and adapt to other kinds of immersive and binding mediums which challenge our human spatial coordinates and ways of being - either down, underwater, or up, in outer space, into networks of vibrant dark matter, and outside the atmospheric critical zone where all known life evolves?
Contemplating the underwater spider using its diving bell not anymore as a water suit allowing her to prey on aquatic insects and crustaceans, but as a spacesuit for exploring not only the atmosphere but also space, is in fact not such a gratuitous analogy. It is no hasard if the training protocol for NASA astronauts involves spending extended periods of time deep below the water’s surface. Before embarking on a journey to its space station, NASA astronauts in training will spend hours at the Neutral Buoyancy Lab, which houses a mockup of the International Space Station 40 feet underwater. They will typically spend six or eight hours at a time in the submerged station, wearing full space suits that weigh more than 350 pounds.




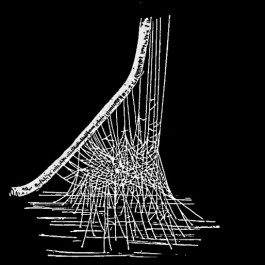
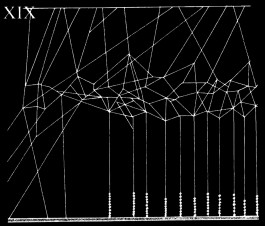
From Henry C. McCook, American spiders and their spinning work. A natural history of the orb-weaving spiders of the United States, with special regard to their industry and habits (1890).
Some species of wasps have evolved to co-opt the central nervous systems of spiders in order to force them to weave stronger webs; in effect, turning the spiders into speculative ‘zombies’. In this scenario, an adult female wasp (Reclinervellus nielseni or Zatypota spp.) lays an egg on the abdomen of a spider (for instance, Cyclosa argenteoalba orAnelosiimus eximius). Once this egg hatches, the wasp larva begins to feed upon the living spider, drinking its hemolymph (blood) and at the same time, injecting substances that will alter the spider’s web-building, and even social, behaviours. In this way, the wasp is able to manipulate the spider to build a particular type of web - a cocoon or ‘resting web’ similar to the web the spider normally builds before molting, but much stronger. Once the spider has completed its task, the larva molts and kills the spider, and then spins the spider’s zombie web into a hanging cocoon, within which the larva will rest and develop for ten days or more.

© Dario J Lagana - Fondazione MAXXI Rome

Collage with Matthäus Merian, 1650 and Jacopo de’ Barbari Map of Venice, 1500.
Sheet webs
Bathyphantes gracilis
Centromerus sylvaticus
Diplocephalus connectens
Erigone dentipalpis
Lepthyphantes istrianus
Lepthyphantes tenuis
Mecopisthes nicaeensis
Meioneta mollis
Meioneta rurestris
Space webs
Amaurobius erberi
Haplodrassus dalmatensis
Microctenonyx subitaneus
Pachygnatha degeeri
Pelecopsis krausi
Prinerigone vagans
Tiso vagans
Trichopterna cito
Steatoda phalerata
Orb webs
Mangora acalypha
Zilla diodia
Tomás Saraceno, Spider/Web map of the Palais de Tokyo, Paris, 2018
© Andrea Rossetti, 2017
Pholcus phalangioides
Eratigena atrica
Amaurobius
From Henry C. McCook, American spiders and their spinning work. A natural history of the orb-weaving spiders of the United States, with special regard to their industry and habits (1890).












